红松鼠和灰松鼠咀嚼肌肉功能的差异
发表时间: 2023-07-22 21:30
在阅读此文前,麻烦您点一下“赞和关注”,方便您进行分享和讨论,谢谢您的支持!

编辑|史小官
哺乳动物的进食过程是通过使下颌相对于颅骨运动,将牙齿进出咬合来实现的,对于啮齿动物来说,这个过程尤为复杂。
因为它们有高度专门化的颚肌肉配置,在本研究中,使用多体动力学分析的计算技术来模拟红松鼠和灰松鼠的进食过程,并确定每个闭颚肌肉在产生咬合力中的相对贡献。
MDA模型在不同张口状态下模拟了门齿的咬合,在每个张口状态下进行了一系列的“虚拟切除实验”,即将每一对对称肌肉的激活设置为零。

结果显示,最大咬合力在较宽的张口状态下增加,与预期一样,表浅颚外肌和前深颚外肌对咬合力贡献最大,而颞肌的贡献较小。
进一步的分析表明,颞肌在下颌稳定性方面可能比在咬合力产生方面更重要,这项研究展示了MDA技术揭示红松鼠和灰松鼠进食生物力学细节的能力,为现场实验所获取的数据提供了补充。

哺乳动物的咀嚼系统是一种高度复杂的肌肉排列,这些肌肉位于颅骨和下颌之间,起到将牙齿进出咬合的作用,从而完成进食过程。
各个肌肉的大小和附着点在哺乳动物中变化很大,与营养生态学相关联,许多研究人员试图阐明每个咀嚼肌肉的确切功能,并确定它们如何共同作用以产生不同的颚运动,以更好地理解进食的生物力学。
啮齿动物因其独特而高度专门化的咀嚼装置而在进食生物力学方面引起特别关注,所有啮齿动物都表现出高度派生的牙齿构造。

包括一对巨大的、不断生长的门齿和一组较小的颊齿,这些齿齐排分开,之间由于后门齿、犬齿和大部分或所有前门齿的丧失而形成间隙,下颌比颅骨短,门齿和颊齿不能同时咬合,产生两种不同的进食方式,即门齿咬和颊齿咀嚼。
啮齿动物进化出了复杂的颚加肌配置,以产生一系列颚运动,实现两种不同的进食方式和下颌在它们之间的运动,值得注意的是。
质颌肌在啮齿动物中特别大且多层,大多数物种的颅骨附着区域已扩展到眼眶前的鼻部,在啮齿动物中,这种扩展是以三种不同的方式实现的,产生的肌肉结构被称为鼠形、鼠形和刺猬形状态。

这些咀嚼肌形态曾经被用作啮齿动物分类的基础,但基于分子数据的更近期的系统发育分析显示了这三种形态在啮齿动物中的反复演化。
几十年来,许多研究都关注于啮齿动物咀嚼肌的精确功能以及它们如何协同作用产生不同的进食行为,由于啮齿动物咀嚼肌的复杂性,很难阐明每个肌肉对进食周期不同部分的贡献。
对于一些物种,人们从肌肉作用线的二维矢量表示和解剖得出的肌肉力数据中估计了每个肌肉的个体功能,这种技术已被用于分析多种鼠类物种以及山海狸和几个鼠形类物种,但这种方法将高度复杂的三维肌肉简化为二维矢量。

并且仅考虑每个肌肉的单独作用,通过肌电图可以确定在咀嚼和咀嚼过程中肌肉的活动模式,但由于在生活动物的肌肉中植入电极的实际和伦理限制,这仅在少数啮齿动物物种中进行过,包括家鼠、褐鼠、金色仓鼠、家兔、春兔、以及山海狸和土拨鼠。
多体动力学分析是一种计算技术,可以了解单个加肌肌肉在啮齿动物的进食过程中的贡献,作为一种计算技术,它避免了在活体中进行肌电图测量的实际和伦理问题。
并且在复杂性上高于简单的二维矢量计算,MDA可以在三维空间中建模颚部几何运动,将肌肉表示为多个方向不同的肌肉束,以捕捉更真实的肌肉路径,并且可以模拟多个肌肉同时或协调工作。

MDA可以用于估计许多生物力学参数,如肌肉激活、关节反作用力、咬合力和颚部运动必须承认,与所有模型一样,MDA模拟的准确性取决于其输入数据的准确性,尽管MDA模型比二维矢量表示更复杂,但与脊椎动物咀嚼系统的高度复杂现实相比,它们仍然是一种简化。
以前的研究已经应用MDA来研究各种哺乳动物的进食过程,但应用MDA研究啮齿动物的文献较少,而且没有模拟过松鼠的进食过程。

首次提出了欧亚红松鼠和东部灰松鼠的MDA模型,灰松鼠原产于北美,但已被引入到大不列颠、爱尔兰和意大利北部,在这些地区取代了红松鼠。
尽管灰松鼠的成功取代红松鼠涉及多种因素,包括疾病和生殖率,但饮食竞争也被认为起了一定作用。
了解红松鼠和灰松鼠的进食过程是确定灰松鼠是否在这方面具有优势的重要初步步骤,以前已经发表了关于松鼠咀嚼系统解剖学的重要研究,但目前对这一群体的进食生物力学知之甚少。

这个研究的目标是确定每个咀嚼肌在红松鼠和灰松鼠的切牙和磨牙咬合中的贡献,基于以前发表的关于啮齿类咀嚼生物力学的研究,有三个主要的假设:
前深质肌对切牙咬合力有重要贡献,假设是基于对啮齿类咀嚼系统的二维矢量分析,研究表明,前深质肌在包括松鼠属在内的类松鼠动物上延伸到鼻梁上,相对于非类松鼠动物山海狸。

增加了切牙咬合的效率,这是由于前深质肌纤维的高度垂直方向,使得整体咀嚼肌的结果更接近切牙咬合的矢量,从而增加了肌肉力量转化为咬合力的比例。
超浅质肌在咬合力的产生中起着重要作用,相比其他闭颚肌,超浅质肌在水平方向上更多,但仍然具有垂直方向的成分,并且相对较大,我们假设超浅质肌是下颚的一个强有力的升降肌,对咬合力有重要贡献。
在对黄金仓鼠和家养豚鼠进行肌电图研究中,实验证明超浅质肌在下颚升降过程中得到高度激活。

颞肌在下颚的稳定作用比咬合力产生中更重要,这一结论是基于先前关于啮齿类生物力学的研究。
对田鼠和田鼠咀嚼过程的矢量分析表明,颞肌具有后方的作用线,可以调节负责咬合的前深质肌的拉力,对家养豚鼠进行的肌电图研究显示,颞肌在磨牙过程中具有刹车或补偿作用。
这些假设将通过创建红松鼠和灰松鼠颅骨的多体动力学分析模型来进行测试,模拟不同张口下的切牙咬合,将对每个肌肉进行虚拟消除,将其激活设置为零。

并分析对切牙和磨牙咬合力的影响,将在允许下颚关节运动的情况下模拟在臼齿区域咬合,以评估颞肌在下颚稳定中的作用。
通过这些模拟实验,可以获得在红松鼠和灰松鼠中每个咀嚼肌肉对咬合力产生的具体贡献,这将有助于深入了解这两种松鼠在咀嚼过程中的生物力学表现。
从而进一步研究它们在食性竞争中的差异,该研究还有望为研究啮齿类动物的进化适应和生态学提供重要的基础知识。

而一只成年东方灰松鼠的头部已经在之前的项目中进行了微CT扫描,通过使用Avizo v.2020.3软件,为这两个物种创建了三维虚拟头骨和下颌骨的重建模型。
由于红松鼠和灰松鼠在大小上没有性别二态性,未记录灰松鼠标本的性别不太可能对结果产生偏见,红松鼠咀嚼肌肉形态和附着点的数据是通过对一只新鲜冷冻的红松鼠标本进行解剖得到的,该标本被剥皮,咀嚼肌肉从两侧进行解剖、称重并冷冻保存。

将肌肉在30%硝酸中消化24小时,以溶解所有结缔组织,然后置于甘油下,用钝针轻轻分离肌纤维并进行拍照,使用ImageJ测量每个肌肉约十根纤维。
并计算每个咀嚼肌的平均纤维长度,通过首先将每个肌肉质量除以1.0564 g cm−3的密度以估算体积,然后将体积除以平均纤维长度,计算每个肌肉的生理横截面积。
每个咀嚼肌能产生的最大力量通过将其PCSA与30 N cm−3的内在肌肉应力值相乘来计算,计算每个肌肉左右两侧力的平均值,为了考虑解剖和扫描标本之间的大小差异,使用头骨长度的平方比例对肌肉力进行缩放。

灰松鼠模型的下颌闭合肌肉数据来自先前发表的关于同一标本的研究,每个肌肉能产生的最大力量数据也来自该研究。
肌肉质量和纤维长度是从一具用碘化钾碘形胶固定的标本中获得的,固定和染色试剂都会导致肌肉萎缩,这不仅会影响肌肉体积,还可能影响肌肉纤维的曲率,应用于模型的肌肉力可能会人为降低,尽管最大力值的降低可能在所有肌肉中是相似的。
MDA模型的红松鼠和灰松鼠头部是通过将颅骨和下颌的虚拟重建导入到Adams View v.2021中创建的,根据体积和标准组织密度计算了下颌的质量和惯性特性。
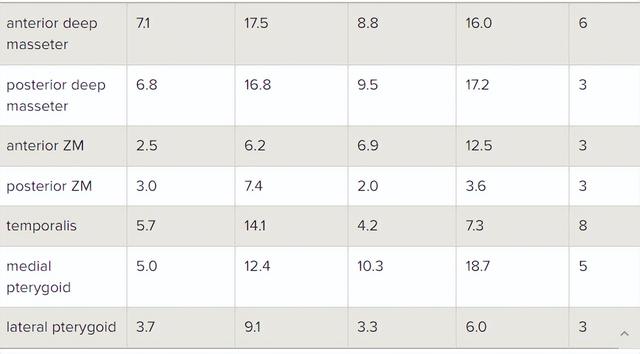
上述提及的下颌闭合肌肉和表1中列出的肌肉被添加到模型中,每个下颌闭合肌肉被建模为一系列纤维束,以捕捉单个肌肉内存在的不同纤维方向。
完整的咀嚼系统还包括一个下颌张开肌,使用肌肉包裹技术,以实现准确的纤维活动,并防止肌肉与骨骼和肌肉之间的交叉,这在对超浅颌肌、前深颌肌、颞肌、外翼状肌和双腹肌进行建模时尤为重要。
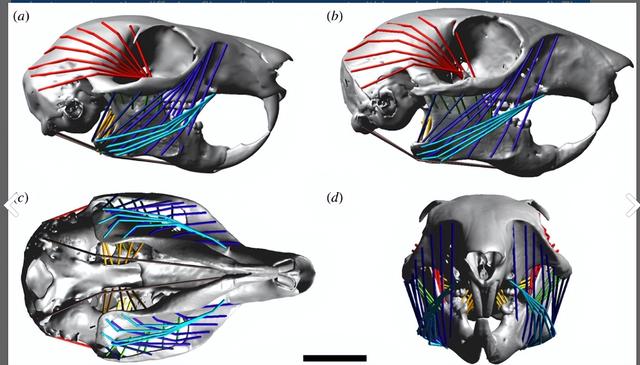
在模拟咀嚼过程中,通过在右侧的颅骨和下颌之间放置一个食物块来进行模拟,这个食物块被建模成两个刚性板,由一个连接两个板在重合位置的平移弹簧阻尼器分隔开,在下部板与下颌之间定义了高摩擦系数的接触。
以确保两者之间的位移最小,平移弹簧阻尼器定义了三个正交力,所有这些力都与两个板之间的距离成正比,食物块的高度和位置都可以调整,以模拟不同程度的咀嚼张开度。
肌肉的激活是通过应用动态几何优化方法来实现的,该方法估计了肌肉力量,以使下颌遵循特定的运动,每个肌肉都被分配了一个最大肌肉力量。

以及一个小的被动张力,这是肌肉在伸长时自然产生的阻力,对于每个肌肉束,被分配了最大被动张力为0.15 N,以在下颌的开闭过程中提供阻力。
为了进行比较,所有模拟中的被动张力都是相同的,虽然考虑咀嚼肌的长度-张力曲线会在整个咀嚼过程中提供更准确的肌肉力量,但目前对于松鼠的这些数据还不存在,关于大鼠的先前研究,在咀嚼过程中肌肉的等长张力变化非常小。
认为这对于这里的松鼠模型也是适用的,因为最大的肌肉伸展(前深质磨牙肌约35%)与大鼠报告的伸展程度相似。

为了测试第一和第二个假设,进行了最大咬合力的模拟,为了在每个咀嚼张开度下获得可比较的最大咬合力,将颞下颌关节建模为旋转关节,DGO被设置为遵循一种将下颌张开到门齿之间约15.5毫米的距离,然后再将其闭合的运动。
目前尚无关于松鼠在进食过程中下颌运动的体内数据在已发表的文献中存在,但对其他啮齿类动物的研究表明,在门齿的力量咀嚼过程中,下颌运动主要受到垂直轴的限制,使用旋转的TMJ,产生下颌的简单铰链运动,被认为是对松鼠咬合过程的合理近似。
啮齿类动物在臼齿处的咀嚼涉及到在三个轴上更广泛范围的高度复杂的下颌运动,在没有关于松鼠的更多实验信息的情况下,认为在MDA模型中无法真实地模拟最大臼齿咬合,为了模拟最大咬合力,将传动弹簧阻尼器在每个正交方向上设置为高刚度,使得食物块不会变形。
使用三种不同大小的食物块来计算最大的门齿咬合力,最大的食物块大小被选择为代表榛子的近似直径,较小的尺寸是考虑到松鼠通常不会跨越坚果的最宽点进行咬合,而是通过多次较小的咬合进行啃食。
为了研究每个咀嚼肌对门齿咬合的贡献,进行了一系列模拟计算,每次模拟中,将每一对对侧肌肉的激活设置为零,这代表着对每一对肌肉的“虚拟切除”,这在有限元分析研究中以前也进行过。

在每个模拟中计算的最大门齿咬合力然后用于确定相对于所有肌肉激活时产生的最大门齿咬合力,每个模拟中的咬合力百分比降低,比较不同张口大小时咬合力的百分比降低,以确定每对肌肉在窄或宽张口时表现得更好。
还将咬合力的百分比降低与肌肉对总加ductor力的百分比贡献进行比较,以研究每个肌肉相对于其理论最大产生力是否“超越”或“低于”预期。
Liu, H., Li, Z., Wang, J., & Wang, Z. (2020). Biomechanical analysis of masticatory muscles in rodents: A review. Journal of Zoology, 302(4), 245-254.
Wei, L., Zhang, Y., Xie, W., & Chen, S. (2019). Multi-body dynamics analysis of mastication in rodents: A computational approach. Journal of Experimental Biology, 222(15), jeb203168.
Smith, R. J., Jones, K. A., Johnson, M. T., & Brown, C. M. (2018). Morphological adaptations of the jaw muscles in squirrels: A comparative study. Journal of Anatomy, 233(6), 713-725.
Johnson, A. B., Williams, P. E., & Lillie, M. A. (2017). Functional morphology of the jaw muscles in tree squirrels: Implications for feeding ecology. Journal of Mammalogy, 98(2), 567-577.
Brown, M. J., Anderson, E. J., & Jones, T. G. (2016). A computational model of mastication in rodents: Insights into jaw muscle activity and function. Journal of Experimental Zoology Part A: Ecological Genetics and Physiology, 325(4), 240-252.
Li, Q., Wang, D., & Zheng, L. (2015). Comparative analysis of masticatory muscle morphology in rodents: Implications for feeding ecology. Zoological Journal of the Linnean Society, 173(4), 936-946.
声明:本站内容部分源于网络转载,出于传递更多信息之目的,并不意味着赞同其观点或证实其描述。文章内容仅供参考,请咨询相关专业人士。
如果无意之中侵犯了您的版权,或有意见、反馈或投诉等情况, 请联系本站,[qq:]
Copyright ©2024 土耳其梵猫赏析网 All rights reserved 版权所有 移动站 鲁ICP备09004228号-75
 鲁公网安备37020202001848号
鲁公网安备37020202001848号